杨维才研究组发现转录抑制机制决定被子植物中央细胞命运
| 杨维才研究组发现转录抑制机制决定被子植物中央细胞命运 |
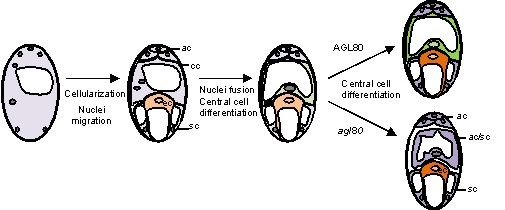
被子植物是当今植物界中、种类最多、分布最广、适应性最强的类群。有别于其它植物类群,被子植物进化出了独特的双受精生殖模式,即雄配子体花粉中的两个精细胞分别与雌配子体内部的卵细胞和中央细胞融合,并进一步发育成胚和胚乳。被子植物双受精机制的出现导致了胚乳的产生,能够为新生的胚提供必要的养分从而确保胚的正常发育,为后代的繁衍和兴盛奠定了基础。中央细胞作为胚乳的前体细胞,其功能和产生的机制一直是植物有性生殖领域研究的热点和难点。 雌配子体——胚囊的发育起始于大孢子母细胞,随后又经过了减数分裂、有丝分裂以及细胞分化等一系列生物学过程最终发育为成熟的雌配子体。大部分被子植物的成熟雌配子体里有四类细胞,分别是位于珠孔端的两个助细胞和一个卵细胞、位于合点端的三个反足细胞和处于胚囊中央位置的中央细胞。其中,除卵细胞外,中央细胞作为第二个雌配子参与调控花粉管吸引和与精子融合,是被子植物所特有的,也是双受精的关键。近些年来,人们对于胚囊里这四类细胞的命运决定机制的研究从未间断,越来越多的调控胚囊发育和细胞分化的基因被鉴定。但对于胚囊内中央细胞的命运决定机制的研究一直很少。 2020年3月4日,分子发育生物学国家重点实验室杨维才研究组在PNAS在线发表了题为“Transcriptional repression specifies the central cell for double fertilization”(DOI:10.1073/pnas.1909465117) 的研究论文。该研究揭示了中央细胞特异的转录抑制复合体AGL80-TPL/TPRs决定中央细胞命运进而影响其受精的分子机制,且进一步发现这一机制在十字花科植物中具有保守性,为人们研究中央细胞的起源及受精过程提供了线索和可能的研究方向。 AGL80是一个在中央细胞特异表达的Type I MADS-box家族的转录因子,前人的研究发现其突变体的中央细胞变小且无法启动胚乳的发育,推测其可能参与了中央细胞的发育过程。在此基础上,本文的研究人员进行了受精实验,发现其agl80中央细胞无法完成受精,而卵细胞的受精不受影响。进一步,研究人员发现突变体的中央细胞表达了助细胞(如MYB98)和反足细胞(如DD1)特异的基因,即agl80中央细胞可能获得了非配子细胞的命运。 通过对AGL80蛋白序列分析,研究人员发现其C端含有一个EAR motif,并发现AGL80能够与转录抑制复合体TOPLESS(TPL)家族五个成员发生互作,该互作依赖于EAR motif。不仅如此,含有EAR motif突变的AGL80不能互补突变体异常的中央细胞。这些结果表明AGL80很可能与TPL家族形成转录抑制复合体在中央细胞的命运决定中发挥功能。为了进一步证明AGL80在胚囊内的转录抑制活性,研究人员将AGL80异位表达在助细胞,结果发现转基因植株出现了不同程度的花粉管导向缺陷表型,并发现参与花粉管导向的助细胞特异基因MYB98的转录呈现不同程度的下调,且表达量与花粉管导向缺陷的表型呈负相关,推测异位表达的AGL80可能抑制了助细胞中MYB98的转录。 鉴于MADS转录因子结合CArG box顺式元件,研究人员首先通过EMSA实验证明AGL80能够结合MYB98启动子区的三个CArG box序列,进而发现突变了CArG box的MYB98可以在中央细胞表达,即启动子区CArG box突变的MYB98在中央细胞的转录不能被抑制。有趣的是,研究人员又进一步将中央细胞特异表达的基因DD22启动子驱动mCherry报告基因转入异位表达AGL80于助细胞的转基因植株,发现在助细胞中也能够检测到红色荧光,说明助细胞异位表达的AGL80能够在助细胞中启动DD22的转录。这些结果表明AGL80-TPL/TPRs复合体不仅能直接抑制助细胞特异基因如MYB98在中央细胞的转录,而且还可能通过抑制某个中间因子间接激活中央细胞特异基因,如DD22,在中央细胞的表达。为了进一步探究这一分子机制在进化上的保守性,研究人员进行了进化和遗传互补分析,AGL80-TPL/TPRs的这一转录抑制机制在十字花科植物中是保守的。
图: AGL80决定中央细胞命运。cc, 中央细胞;ec, 卵细胞;sc, 助细胞;
分子发育生物学国家重点实验室李红菊研究员和杨维才研究员为该论文的共同通讯作者,已毕业的张梦夏博士和朱杉杉博士为该论文的共同第一作者。中科院植物所郭亚龙研究员和博士生徐永超也参与了部分工作。本论文得到国家自然科学基金等的支持。 |
附件下载: